Adam Pines

My work centers on delineating the development of the brain, cognition, and psychopathology with data science. Rather than judging where our brains and minds may be at any one point in time, I believe our trajectories define us. By understanding how disorders emerge over time, we may provide the means to understand how they remit.
“Every science begins as philosophy and ends as art” - Durant
CV ||| github ||| google scholar ||| twitter ||| bluesky
Publications
First-author
Psychedelics disrupt hierarchical cortical propagations in the default mode network of humans and mice
Pines, Zhang, Kochalka … & Williams, 2026
Psychiatric Symptoms, Cognition, and Symptom Severity in Children
Pines, Tozzi, Bertrand … & Williams, 2024
Development of Top-Down Cortical Propagations in Youth
Pines, Keller, Larsen … & Satterthwaite, 2023
Dissociable multi-scale patterns of development in personalized brain networks
Pines, Larsen, Cui … & Satterthwaite, 2022
Leveraging multi-shell diffusion for studies of brain development in youth and young adulthood
Pines, Cieslak, Larsen … & Satterthwaite, 2020
Multi-unit relations among neural, self-report, and behavioral correlates of emotion regulation in comorbid depression and obesity
Pines, Sacchet, Kullar …. & Williams, 2018
Second and Third Author
Adaptive cognitive control circuit changes associated with problem-solving ability and depression symptom outcomes over 24 months
Zhang, Pines, Stetz … & Williams, 2024
Individual Differences in Delay Discounting are Associated with Dorsal Prefrontal Cortex Connectivity in children, adolescents, and adults
Mehta, Pines, Adebimpe … & Satterthwaite, 2023
Linking Individual Differences in Personalized Functional Network Topography to Psychopathology in Youth
Cui, Pines, Larsen … & Satterthwaite, 2022
Personalized Functional Brain Network Topography is Associated with Individual Differences in Youth Cognition
Keller, Pines, Sydnor … & Satterthwaite, 2023
The ENGAGE study: Integrating neuroimaging, virtual reality and smartphone sensing to understand self-regulation for managing depression and obesity in a precision medicine model
Williams, Pines, Rosas … & Ma, 2018
Personalized brain circuit scores characterize depression biotypes with distinct symptoms, behavioral profiles, and treatment outcomes
Tozzi, Zhang, Pines … & Williams, 2024
Functional Connectivity Development along the Sensorimotor-Association Axis Enhances the Cortical Hierarchy
Luo, Sydnor, Pines … & Satterthwaite
Pairwise maximum entropy model explains the role of white matter structure in shaping emergent co-activation states
Ashourvan, Shah, Pines … & Litt, 2021
Associations between neighborhood socioeconomic status, parental education, and executive system activation in youth
Murtha, Larsen, Pines … & Satterthwaite, 2022
Caregiver monitoring, but not caregiver warmth, is associated with general cognition in two large sub-samples of youth
Keller, Mackey, Pines … & Satterhtwaite, 2022
Hierarchical functional system development supports executive function
Keller, Sydnor, Pines … & Satterthwaite, 2022
Mapping the Neurodevelopmental Predictors of Psychopathology
Jirsaraie, Gatavins, Pines … & Sotiras, 2024
A public database of immersive VR videos with corresponding ratings of arousal, valence, and correlations between head movements and self report measures
Li, Bailenson, Pines … & Williams, 2017
Middle Author
QSIPrep: An integrative platform for preprocessing and reconstructing diffusion MRI
Cieslak, Cook, He … & Satterthwaite, 2022
A Precision Functional Atlas of Network Probabilities and Individual-Specific Network Topography
Hermosillo, Moore, Fezcko … & Fair
Neurodevelopment of the association cortices: patterns, mechanisms, and implications for psychopathology
Sydnor, Larsen, Bassett, … & Satterthwaite, 2021
A Developmental Reduction of the Excitation:Inhibition Ratio in Association Cortex during Adolescence
Larsen, Cui, Adebimpe … & Satterthwaite, 2022
Deciphering the functional specialization of whole-brain spatiomolecular gradients in the adult brain
Vogel, Alexander-Bloch, Wagstyl … & Seidlitz
Characterizing the role of the structural connectome in seizure dynamics
Shah, Ashourvan, Mikhail … & Davis, 2019
Sex differences in functional topography of association networks
Shanmugan, Seidlitz, Cui … & Satterthwaite, 2021
A general exposome factor explains individual differences in functional brain network topography and cognition in youth
Keller, Moore, Luo … & Barzilay
Mobile Footprinting: Linking Individual Distinctiveness in Mobility Patterns to Mood, Sleep, and Brain Functional Connectivity
Xia, Barnett, Tapera … & Satterthwaite, 2022
Developmental coupling of cerebral blood flow and fMRI fluctuations in youth
Baller, Valcarcel, Adebimpe … & Satterthwaite, 2022
An analysis-ready and quality controlled resource for pediatric brain white-matter research
Richie-Halford, Cieslak, Ai … & Rokem, 2022
Preprints
Compression supports low-dimensional representations of behavior across neural circuits
Zhou, Kim, Pines … & Bassett
Connectional Hierarchy in Human Brain Revealed by Individual Variability of Functional Network Edges
Yang, Wu, Li … & Cui
About me
I’m a postdoctoral scholar with Anish Mitra and Nolan Williams at Stanford researching how neuropsychopathology emerges and remits the developing brain. I completed my neuroscience PhD at the University of Pennsylvania with Ted Satterthwaite, where I studied normative neurocognitive development.
My goal is to build the neuroscientific foundations needed to advance mental health care, and in parallel, to translate this research to meaningful real-world outcomes. I tend to focus on the interplay between mood disorders and cognitive dysfunctions, with specific expertise in how depression and cognitive functions emerge in adolescence. Neuroscientifically, many of these advances are rooted in hierarchical cortical organization and hierarchical cortical reorganization in development.
The movement of the scientific community towards transparency is more important than any work within one specific subfield. As I’ve relied on previous generations of open-source science, I’ve committed to this movement by creating walkthroughs for all of my code underlying every analysis in my first-author papers since learning how to code. All of the code within these walkthroughs (and the walkthroughs themselves) have been independently verified to work as intended by particularly stalwart co-authors. For example, in this pre-print, we were able to build off of fantastic code bases for spherical registration of cortical surfaces (freesurfer) and for tracing the migration of progenitor cells on spherical zebrafish gastrulas to construct, validate, hypothesis-test with, and disseminate a pipeline for tracking and quantifying the hierarchical movement of cortical activity.
Outside of neuroscience, I’m a long-time mediocre improvisational musician, enjoy backpacking into the mountains with my wife and friends, am an ex-personal trainer, and am active in the great American folk science of meat smoking. Thanks for visiting my page, and feel free to reach out for inquiries. Here are some findings I’d like to highlight:
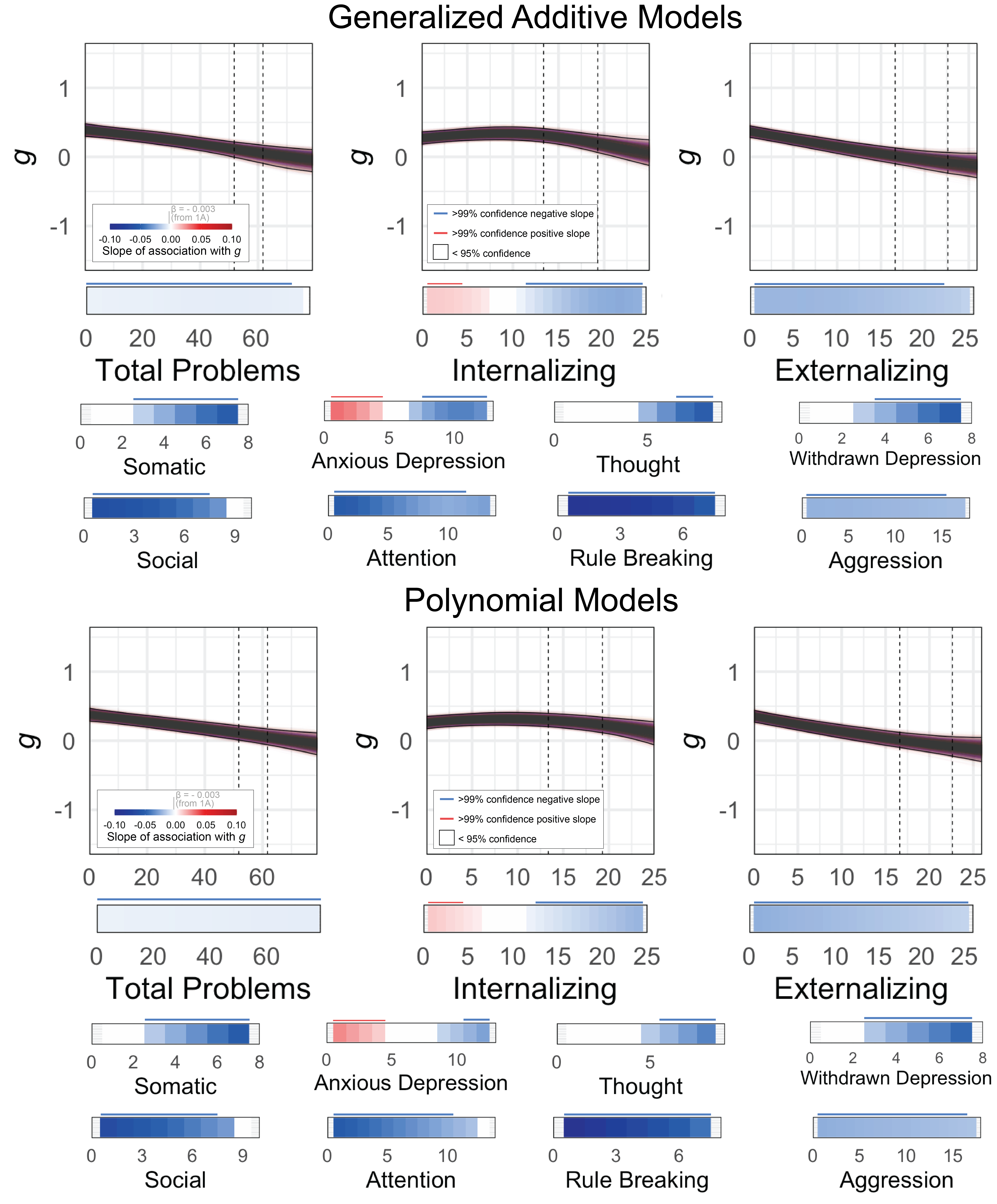
Whether cognition is positively or negatively associated with symptoms depends on the class and magnitude of symptoms. Positive and negative relationships are robust to bootstrapping across modeling techniques at >99% statistical confidence, but co-exist within the same class of symptoms. These results can explain dozens of discrepant papers, often with strong methods and large sample sizes, arguing that cognition is either negative or positively associated with depression symptoms. If you look more closely at these studies, you’ll see the papers arguing for a positive association (higher cognitive scores = higher symptom scores) are almost all in subclinical samples, whereas papers arguing for negative associations (higher cognitive scores = lower symptom scores) are almost all in clinical samples. Both sets of papers are “correct”, but relying on linear models in samples that did not cover the full spectrum of mental health obscured that the association between cognition and internalizing symptoms depends on how well or unwell an individual is. Dashed lines represent borderline and clinical thresholds from the CBCL. From my JAMA psych paper.
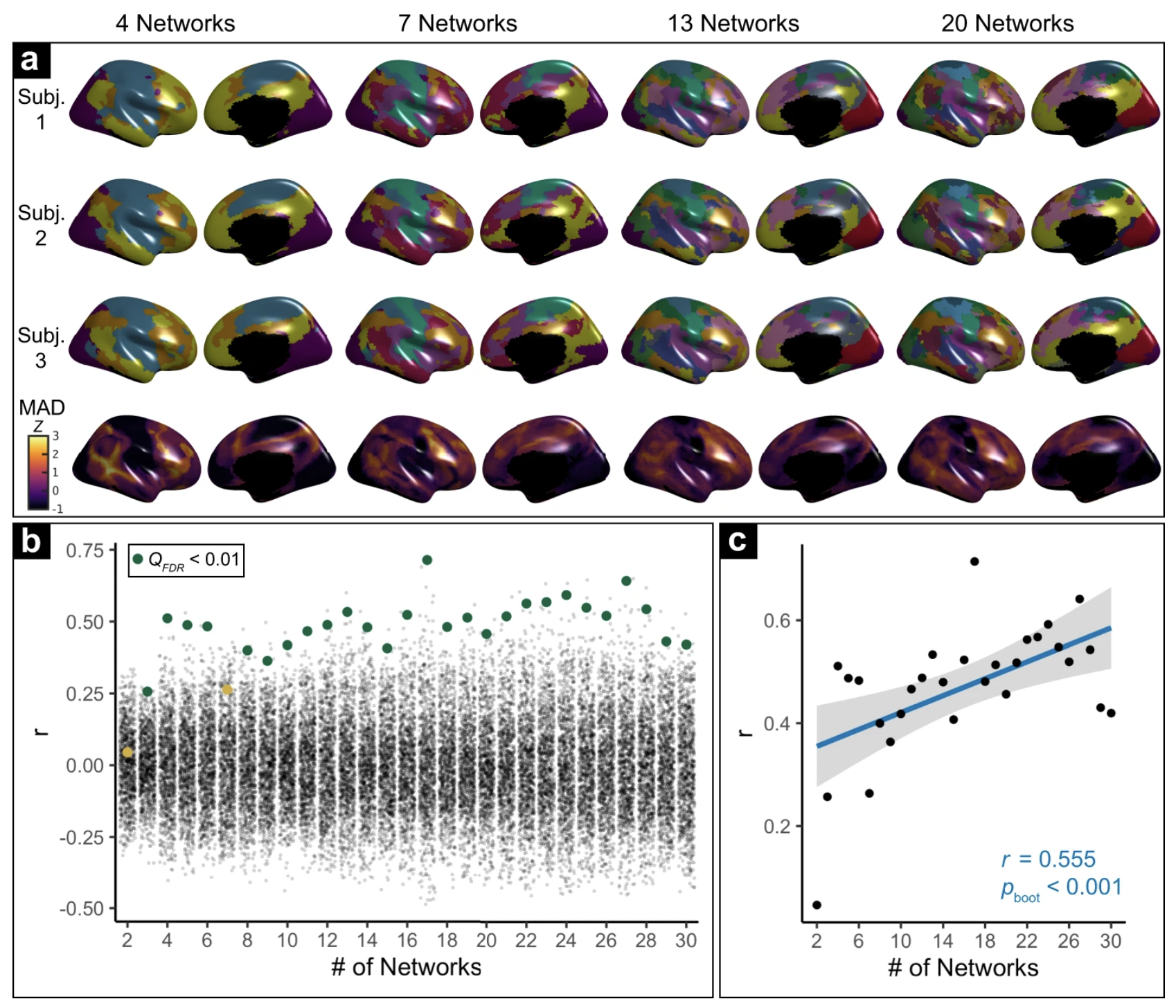
Where in the cortex individual variability in functional arealization localizes is dependent on how granular your definition of functional networks is. More granular = more variability in higher-order cortices. From my nature comms paper.
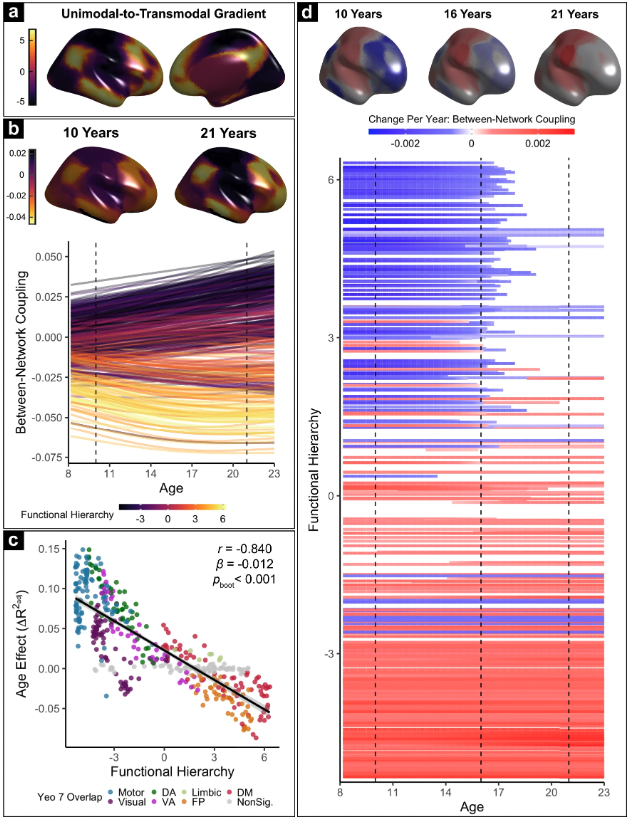
Functional differentiation proceeds from lower-order to higher-order cortices in development, such that a single unimodal-to-transmodal gradient explains most (r^2=.71) of the developmental variability we observed across functional networks. Functional differentiation is associated with enhanced cognitive capacity, and de-differentiation seems to follow the same spatial trajectory years-to-decades later (still waiting to run this on full lifespan data). From my nature comms paper.
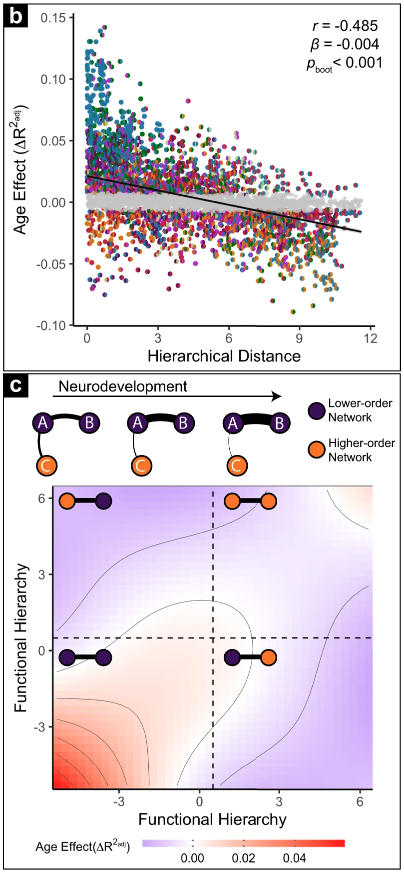
Hierarchical distance provides a parsimonious description of how individual edges in the functional connectome develop. This pattern has since been replicated in 4 independent datasets (Luo et al., 2024, Nat. Comms). Check out figure S9 from my paper (also Nat. Comms, 2022) to see how it lines up relative to other edge-level descriptions of neurodevelopment.
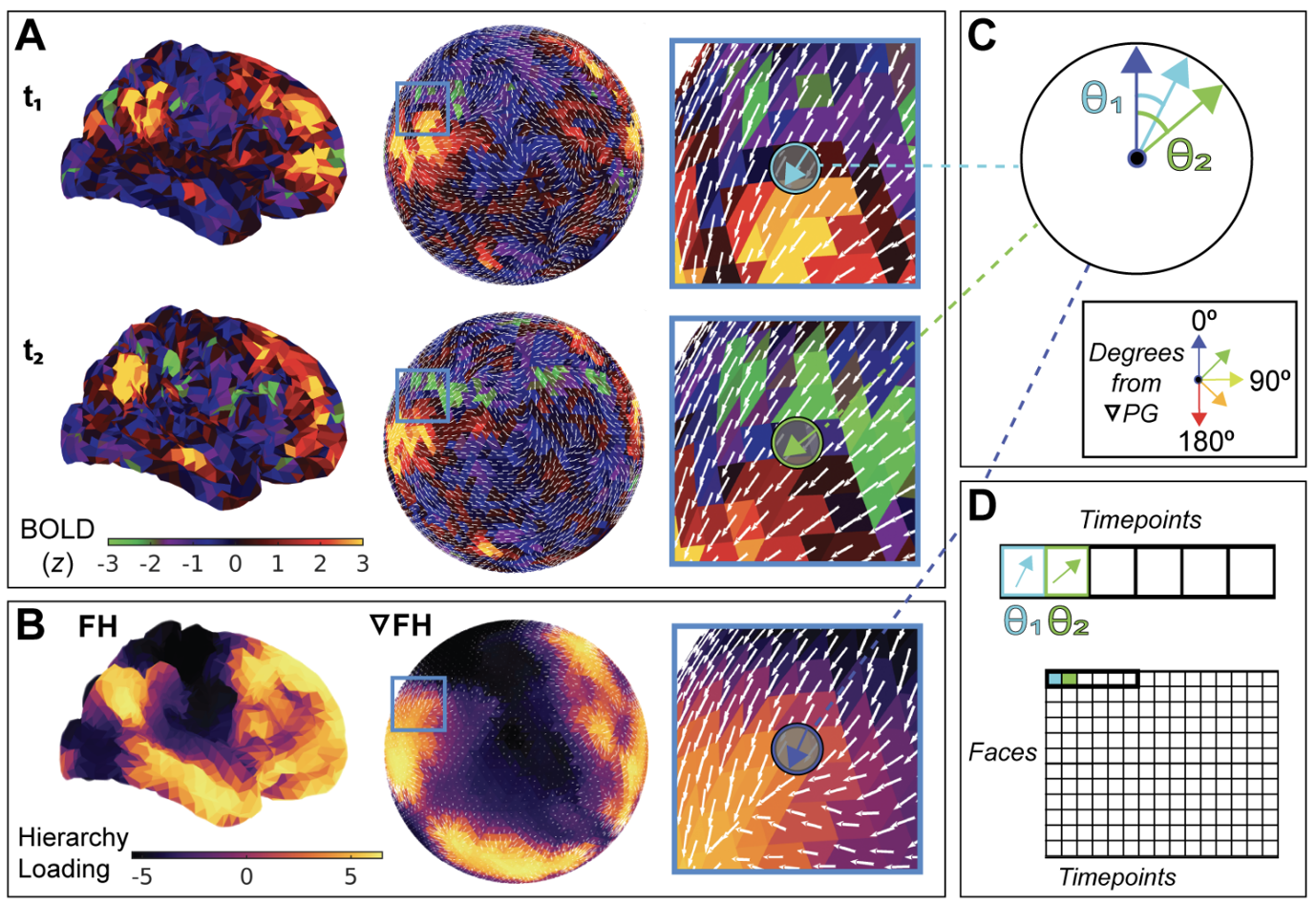
Optical flow can be used to delineate bottom-up and top-down hierarchical propagations in fMRI data. From my neuron paper.
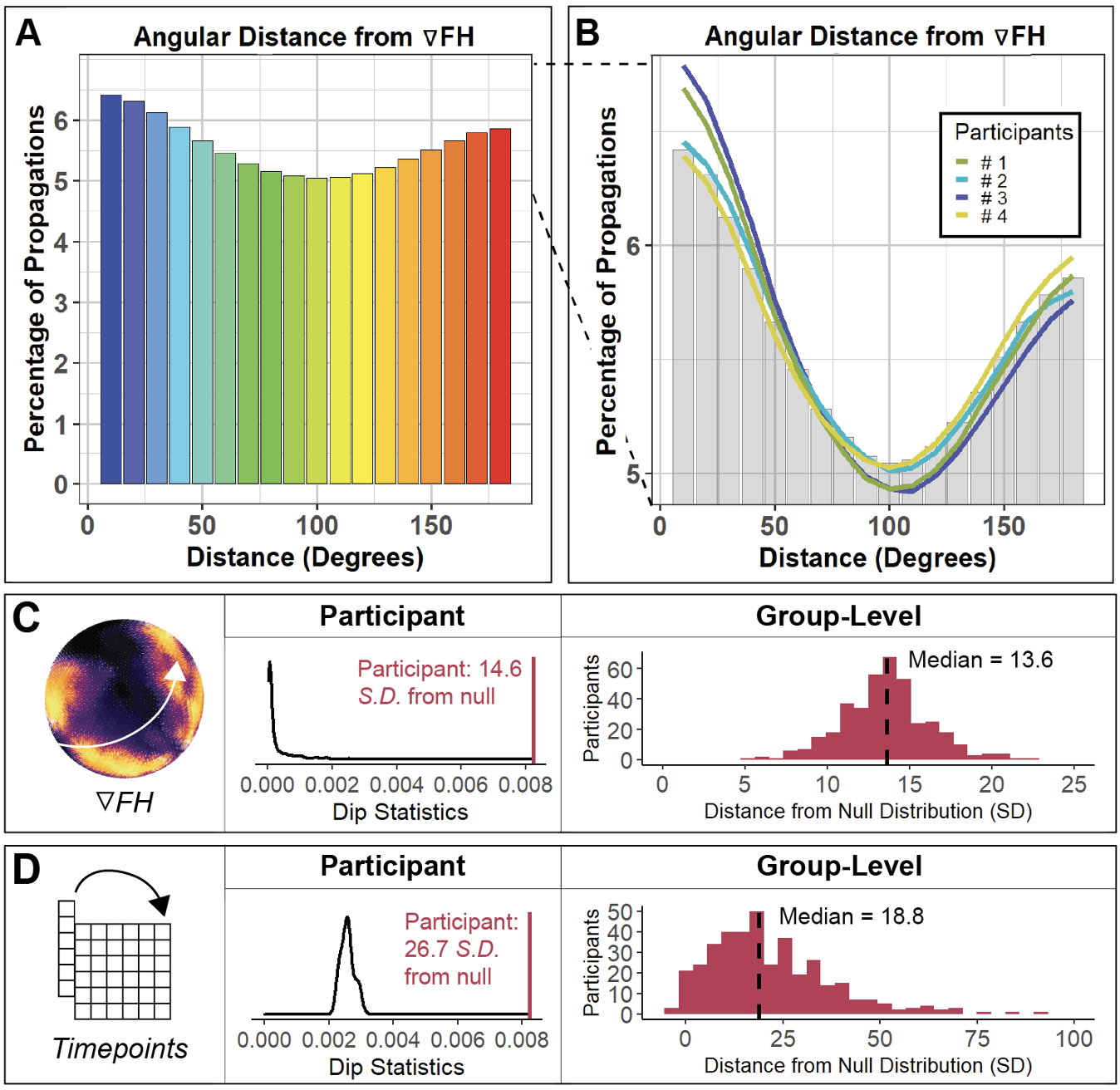
Both bottom-up and top-down propagations are common in all individuals we tested. 2 of 5 verifications of this finding depicted in this figure. From my neuron paper.
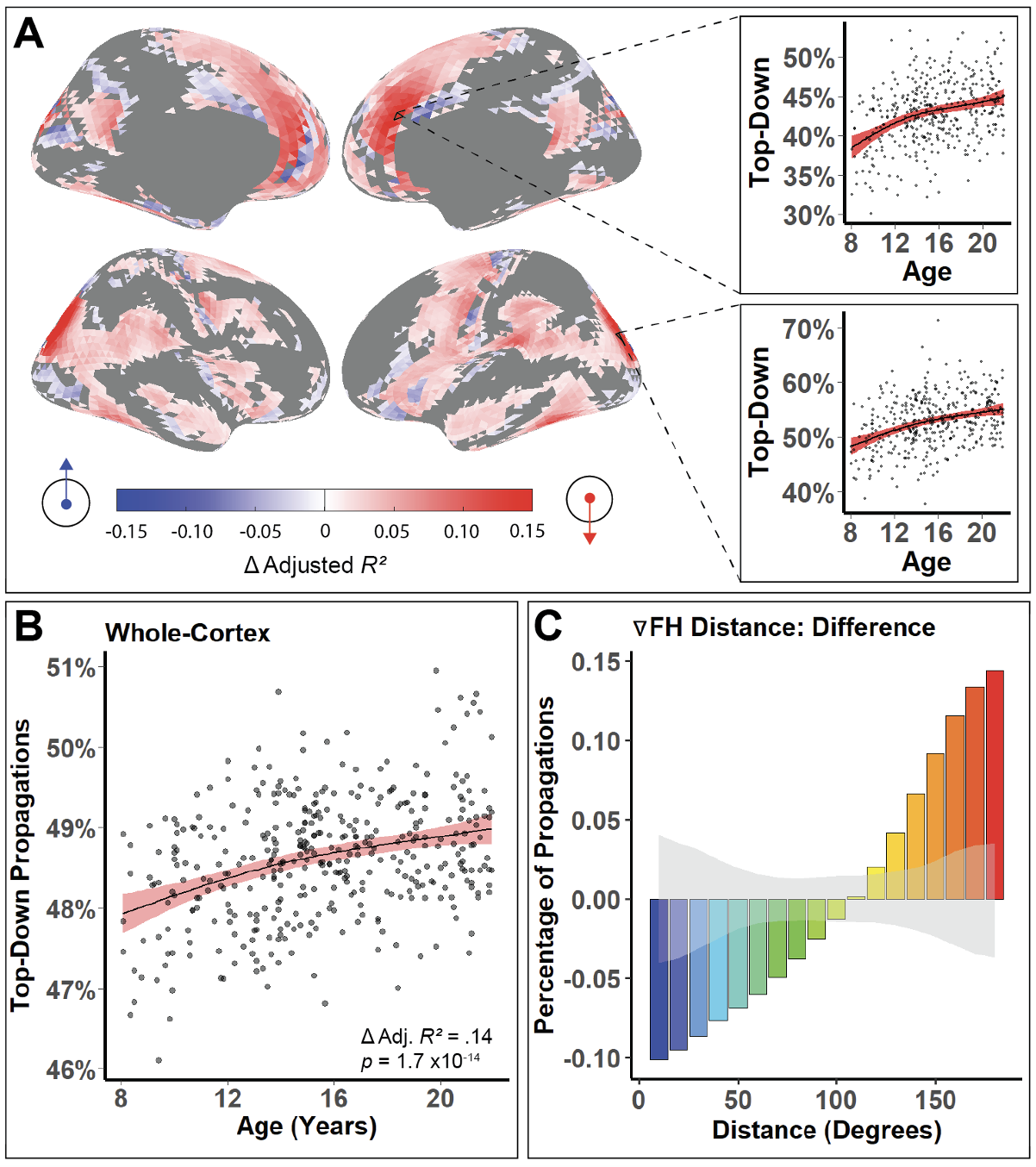
Top-down propagations become increasingly prominent over neurodevelopment. Not pictured is the same results 100% holding after controlling for previously known properties of functional neurodevelopment. From my neuron paper.
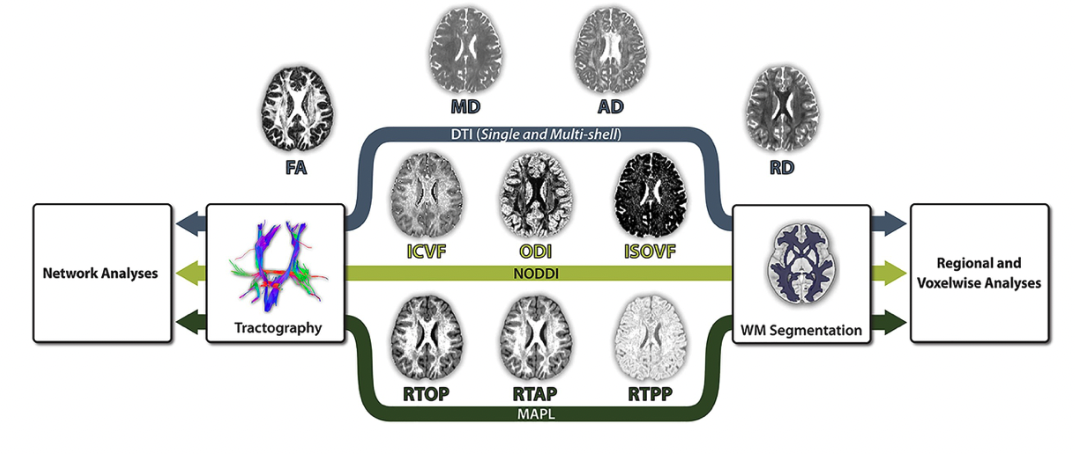
Multi-shell diffusion weighted imaging can be leveraged to confer increased sensitivity to neurodevelopmental effects and decreased sensitivity to the confounding influence of head motion in neurodevelopmental studies. Schematic from my developmental cog. neurosci. paper.